Sain alla olevan vieraskirjoituksen jälleen tuttavaltani. Hänellä on korkein mahdollinen alan koulutus ja yli 30 vuoden kokemus lääketeollisuudessa työskentelemisestä ja lääketieteellisestä tutkimustyöstä. Nykyisessä sensuurin ja totalitarismin ajassa hän haluaa pysyä anonyyminä. Jo aiempi kirjoitus oli kristallinkirkkaan ajattelun tulos – siinä on 28 viitettä ja todellisen asiantuntijan synteesi. Tässä kirjoituksessa korruptiopandemian tilanne päivitetään – 122 viitettä! (OT)
Kirjoitin elokuussa 2021 kuinka maailmaa vaivasi maaliskuusta 2020 lähtien koronapandemian sijasta korruptiopandemia, jossa tähtitieteellisiä voittoja tekevä rokoteteollisuus korruptoi poliitikot, terveysviranomaiset, lääkevalvontaviranomaiset, johtavat asiantuntijat ja valtamedian sensuroimaan rokotehaittoja ja suoltamaan tiedevastaisia valheita, joiden tarkoituksena oli maailman väestön orjuuttaminen rokoteteollisuuden kanta-asiakkaiksi. Käyn tässä kirjoituksessa läpi mitä korruptiopandemiasta nyt tiedetään, ja kuinka tämän kaikkien aikojen törkeimmän rikoksen mahdollistaneet tahot tullaan saattamaan lähivuosina laajalla rintamalla maailmanlaajuisesti rikosoikeudelliseen vastuuseen.
Aluksi lyhyt kertaus valekoronapandemiasta ja miten se toteutettiin. Meille väitetään, että ihmiskunnalle uusi SARS-CoV-2 koronavirus on aiheuttanut poikkeuksellisen tappavan pandemian, ja että virusta vastaan kehitettiin ennätysajassa rokotteita, jotka ovat tehokkaita ja turvallisia. Kuten tästä kirjoituksesta selviää, mikään edellämainituista ei pidä paikkaansa. Mitään yleisvaarallista koronapandemiaa ei ole koskaan ollut, joten millekään koronatoimille kuten maskisuosituksille, yhteiskunnan rajoitustoimille, koronarokotuksille ja koronapassille ei ole koskaan ollut mitään lääketieteellisiä perusteita. Koronarokotteet ovat tarpeettomia, tehottomia, terveydelle haitallisia ja hengenvaarallisia, ja koronatoimilla on täysin turhaan oikeudettomasti puututtu ihmisten elämään, rikottu räikeästi ihmisoikeuksia ja perustuslakia, ja aiheutettu valtava määrä haittaa kansantaloudelle ja kansanterveydelle.
SARS-CoV-2 viruksen poikkeuksellista vaarallisuutta perusteltiin valepandemian alussa sillä, että virus olisi ihmisille täysin uudenlainen, minkä vuoksi ihmisillä ei olisi sille minkäänlaista vastustuskykyä. Tämä ei kuitenkaan pidä paikkaansa. Useita tavallista flunssaa aiheuttavia koronaviruksia on kiertänyt vuosikymmeniä (viite 1), ja SARS-CoV-2 virus on niiden kanssa riittävän samankaltainen, minkä vuoksi iso osa ihmisistä on ollut sille ennestään vastustuskykyisiä (viite 2). Viruksen sekvenssi on myös löydetty yli 120:sta viimeisen reilun 20 vuoden ajan julkaistusta patentista (viite 3, viite 4), joten se on ollut tunnettu jo kauan ennen valepandemiaa. Patenttitiedot viittaavat myös vahvasti siihen, että SARS-CoV-2 -virus on kehitetty laboratoriossa, ja onpa myös vahvasti kyseenalaistettu onko virusta edes olemassa muualla kuin tietokoneen ruudulla (viite 5, viite 6).
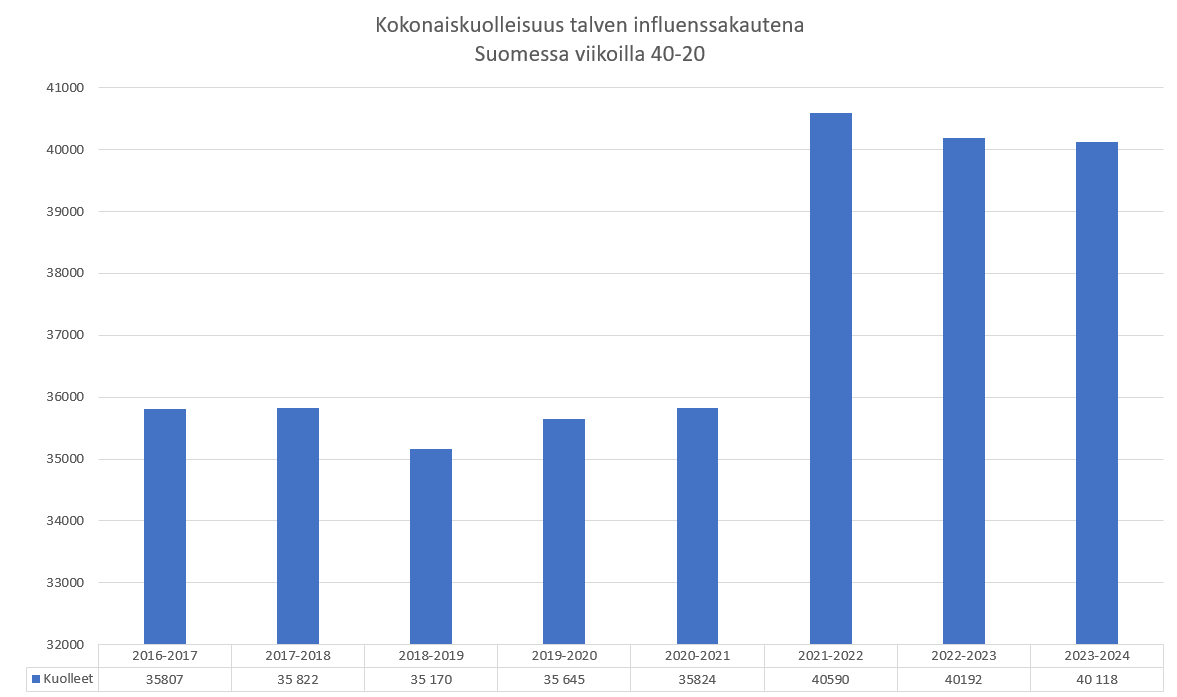
Stanfordin yliopiston arvostetun Prof. John Ioannidisin loppuvuonna 2021 tekemän analyysin mukaan korona oli vuonna 2020 vaarallisuudessa verrattavissa tavalliseen flunssaan (viite 7). Koronan vaarallisuutta liioiteltiin vahvasti myös Suomessa, sillä vuonna 2020 koronaan kuoli virallisten tilastojen mukaan ainoastaan 558 henkilöä, joista lähes kaikki olivat monisairaita vanhuksia, ja kuolleiden keski-ikä oli 84 vuotta (viite 8). Vuonna 2020 korona ei myöskään lisännyt kokonaiskuolleisuutta (viite 9), eikä se ole missään vaiheessa täyttänyt lähimainkaan Sosiaali- ja Terveysministeriön luokituksen mukaisia pandemian kriteerejä (viite 10). Ns. ”vakavan koronataudin” tärkeimmiksi riskitekijöiksi on paljastunut ylipaino (viite 11), ja veren alhainen D-vitamiinitaso (viite 12).
Koronadiagnoosit perustuivat varsinkin valepandemian alkuaikoina PCR-testiin. PCR-teknologian kehittäjä, Nobel-palkittu Kary Mullis on todennut, että PCR-testi ei sovellu diagnostiseen käyttöön (viite 13). SARS-CoV-2 PCR-testissä on useita epäkohtia, kuten se, että testi tunnistaa kaikki vuosikymmeniä kiertäneet tavallista flunssaa aiheuttavat koronavirukset sekä niiden palaset (viite 14, sivut 18-19). Testiä myös tehtiin Suomessakin yleisesti 40-45 monistuskerralla, vaikka monistuskertojen määrä ei saisi olla yli 24 (viite 15), ja jo 35 monistuskerran jälkeen peräti 97% positiivisista tuloksista on vääriä (viite 16). Väärien positiivisten koronadiagnoosien seurauksista oltiin huolissaan jo syksyllä 2020 Lancet-tiedelehdessä julkaistussa tutkimuksessa (viite 17), ja WHO huomautti asiasta joulukuussa 2020 (viite 18). Nämä huolet kaikuivat kuitenkin koronahuumassa/pelossa kuuroille korville.
PCR-testejä tehtiin paljon myös oireettomille, ja heitä sekä heille ”altistuneita” määrättiin positiivisten testitulosten seurauksena karanteeniin. Tämä siitä huolimatta että jo vuosikymmenten ajan on tiedetty, että oireettomat eivät voi tartuttaa hengitystieviruksia, mikä on todistettu myös SARS-CoV-2 viruksen osalta laajoissa tutkimuksissa (viite 19, viite 20). Oireettomia on myös vaadittu käyttämään kasvomaskeja, vaikka edes parhaat kirurgiset maskit eivät suojaa virusinfektioilta sen parammin käyttäjiään kuin muita, vaan kasvomaskien pitkäaikaiskäyttö voi aiheuttaa vakavia terveyshaittoja (viite 21, viite 22, viite 23). Myös käsidesin liiallinen käyttö on terveydelle haitallista (viite 24), ja millään yhteiskunnan rajoitustoimilla ei voida ehkäistä virusinfektioiden leviämistä (viite 25).
Valtaosalla koronavirusinfektion saaneista oireet ovat lieviä ja verrattavissa tavalliseen flunssaan. Muiden flunssavirusten tavoin koronavirukset voivat aiheuttaa vakavan taudin erityisesti monisairaille vanhuksille, sekä myös muille joiden immuunipuolustus on heikentynyt. Hengitystieinfektioiden aiheuttaman vakavan taudin tehokas ja turvallinen ehkäiseminen ja hoito voidaan toteuttaa esim. seuraavasti:
- Ehkäiseminen vahvistamalla luontaista immuniteettia D-vitamiinilla (viite 26, viite 27).
- Oireiden ilmaannuttua varhaiseen hoitoon on olemassa erittäin tehokkaita ja turvallisia lääkkeitä, kuten Nobelpalkittu ivermektiini, joka kuuluu WHO:n maailman tärkeimpien lääkkeiden listaan, ja jonka avulla korona on selätetty täydellisesti useissa maissa (viite 28, viite 29). Ivermektiinin hoito-ohjeita on julkaistu myös Suomeksi (viite 30).
- Vakava koronatauti voidaan hoitaa tehokkaasti steroideilla (viite 31, viite 32).
Sairaaloille maksettiin koronapotilaista merkittäviä rahallisia korvauksia, ja sairaalat myös ohjeistettiin tekemään PCR-testejä liian suurilla monistuskerroilla, jolloin useat muutkin hengitystieinfektiot diagnosoitiin väärin koronaksi, minkä seurauksena mm. influenssa katosi salaperäisesti maailmasta ”koronapandemian” alettua. Suomessakin valtion sairaanhoitopiireille jakamien korona-avustusten määrä liitettiin koronapotilaiden toteutuneiden hoitovuorokausien määrään (viite 33). Yhdysvalloissa (viite 34) sairaalat tehtailivat rahallisten kannustimien houkuttelemina potilailleen ”koronakuolemia” saattamalla heidät tehohoitokuntoon vakavia munuais- ja maksavaurioita aiheuttavalla remdesivirillä (viite 35, viite 36) ja aiheuttamalla heille sitten keuhkokuumeen laittamalla heidät hengityskoneeseen, mihin potilaat lopulta kuolivat (viite 37). Yhdysvalloissa lääkärit saattoivat tienata koronarokotteilla useita satoja tuhansia ylimääräisiä tuloja (viite 38, viite 39).
Suomessakin toteutettiin massiivinen kampanja, joka tähtäsi ”koronarokotteiden” antamiseen koko väestölle, pääasiassa Pfizer-BioNTechin ns. mRNA-valmisteilla, vaikka niiden tiedettiin olevan käytännössä täysin tehottomia sekä Pfizerin omissa tieteellisissä tutkimuksissa (viite 40, viite 41) että laajoissa väestöanalyyseissä (viite 42). Lopulta valmisteiden väitettiin ainoastaan vähentävän vakavia tautimuotoja ja kuolemia, mutta tästäkään ei ole olemassa mitään tieteellistä näyttöä. Niiden kliinisissä tutkimuksissakin selvitetään ainoastaan vähentävätkö valmisteet oireellisten PCR-diagnosoitujen koronatartuntojen määrää. Vakava tautimuoto on niin harvinainen, että tutkimusaineiston koko ei riitä selvittämään, onko valmisteilla vaikutusta siihen (viite 43). Kaikesta huolimatta esim. THL suosittelee valmisteita edelleen erityisesti vanhuksille ja immuunipuutteisille, joilla sen tehoa ja turvallisuutta ei ole edes tutkittu.
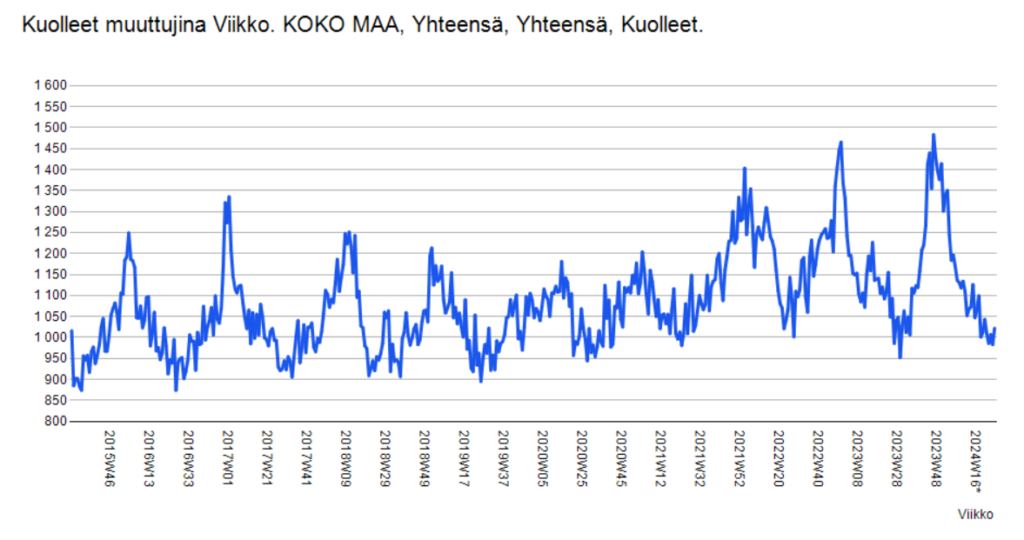
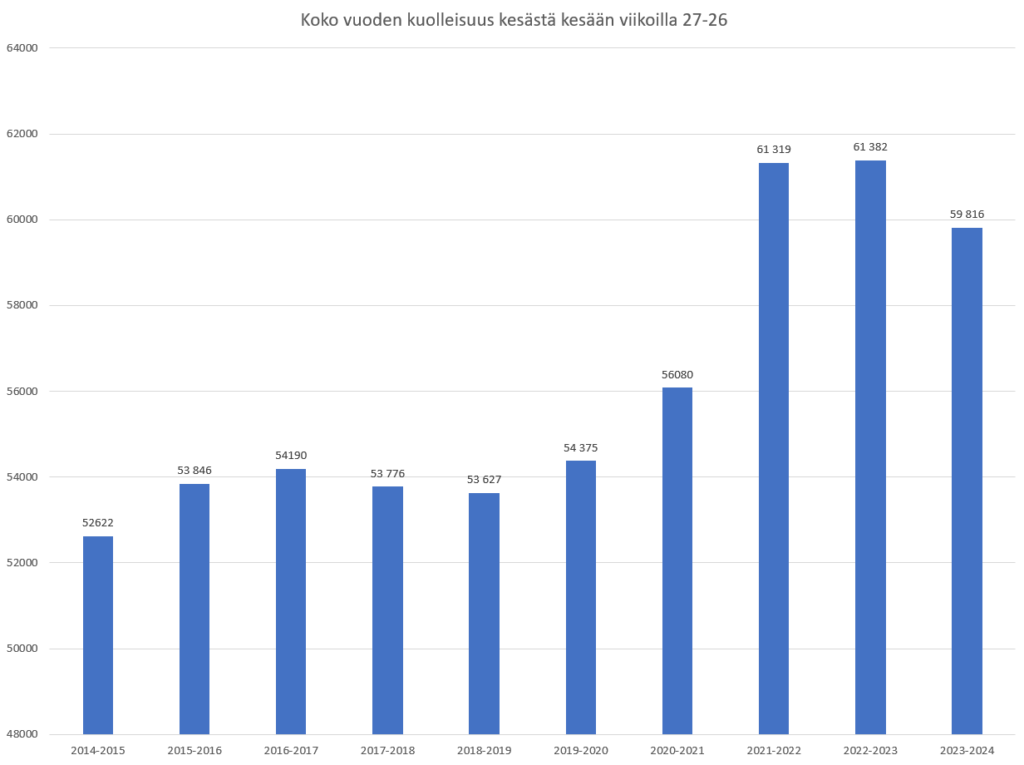
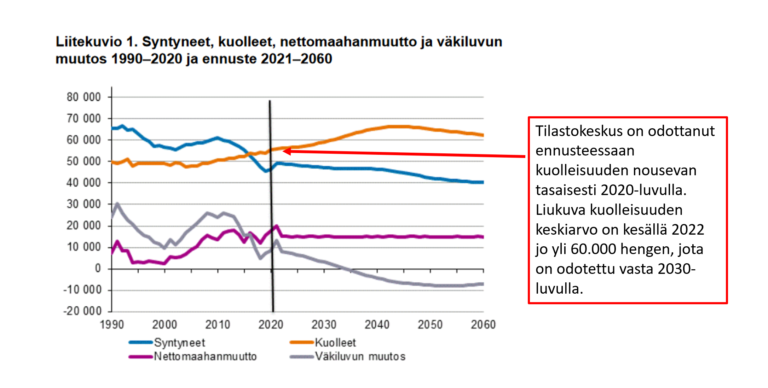
Olemattoman tehokkuuden lisäksi erityisesti mRNA-valmisteiden tiedetään vaikutusmekanisminsa perusteella aiheuttavan valtavan määrän erilaisia terveyshaittoja, jotka on tarkasti dokumentoitu (viite 44), joista oli jo tammikuussa 2022 julkaistu yli tuhat tieteellistä julkaisua (viite 45), ja jotka näkyvät laajalti eri maiden rokotehaittarekistereissä (viite 46) sekä koronapiikitysten aloittamisen jälkeen alkaneena merkittävänä ylikuolleisuutena (viite 47, viite 48). Viitteen 47 yhteenvedossa sivulla 316 todetaan yksiselitteisesti, että korona-aikainen valtava ylikuolleisuus johtui yksinomaan eri valtioiden hallitusten toteuttamista koronatoimista ja terveydelle haitallisista koronarokotteista, ja jos mitään pandemiaa ei olisi julistettu ja rajoituksia ym koronatoimia sekä rokotuksia ei olisi toteutettu, ylikuolleisuudelta olisi kokonaan vältytty. Ylikuolleisuuden lisäksi maailmalla havaittiin laajamittainen syntyvyyden lasku, jonka alkaminen ajoittui noin 9 kk koronarokotusten aloittamisen jälkeen (viite 49, viite 50). Tämä viittaa vahvasti siihen, että koronarokotteet lisäävät paitsi kuolleisuutta myös keskenmenoja.
Koronarokotteiden ja ylikuolleisuuden välinen yhteys on näkynyt myös Suomessa niin selvästi, että kukaan asiaan perehtynyt ei voi olla sitä ymmärtämättä. Esimerkkinä tästä THL suositti syksyllä 2023 uutta koronarokotuskierrosta iäkkäille ja immuunipuutteisille (viite 51). Rokotukset toteutettiin loka-marraskuussa 2023 (viite 52). Rokotusten seurauksena uutisoitiin marraskuun 2023 lopussa, että syksyn korona-aalto osui iäkkäisiin voimakkaammin kuin koskaan ennen (viite 53), ja myös tilastokeskus uutisoi joulukuussa 2023, että Suomessa kuoli kyseisen rokotuskierroksen jälkeen ennätysmäärä ihmisiä (viite 54). Julkisuudessa esitetyt väitteet siitä, että syynä olisi ollut rokotusten viivästyminen, ovat epätoivoinen yritys peitellä ilmiselviä rokotekuolemia.
Pfizerin ja Modernan ”mRNA-rokotteet” eivät itse asiassa sisällä lainkaan mRNA:ta vaan lipidinanopartikkeleihin pakattua ns. modRNA:ta (mod. tulee sanasta ’modified’ eli muokattu), jossa luonnollisessa RNA:ssa esiintyvä urasiili on korvattu pseudouridiinilla, mikä johtaa molekyylin vapaaseen kulkeutumiseen ympäri kehoa, pitkäaikaiseen stabiloitumiseen ja useisiin terveyshaittoihin kuten luonnollisen immuniteetin heikentymiseen ja syöpäriskin lisääntymiseen rokotetuilla (viite 55, viite 56). Huolestuttavaa on myös se, että ”mRNA-rokotteiden” on todistettu sisältävän huomattavan määrän mm. syövälle altistavaa DNA:ta epäpuhtautena (viite 57, viite 58). Valmisteiden on myös osoitettu voivan päätyä osaksi solujen DNA:ta (viite 59), joten niiden haitat saattavat ulottua myös seuraaviin sukupolviin. Rokotteiden sisältämät nanopartikkelit on tarkoitettu ainoastaan tutkimuskäyttöön, ja niitä ei saa antaa ihmisille, mutta siitä huolimatta niitä sisältäviä valmisteita on annettu miljardeille ihmisille (viite 60).
Pfizer ja sen mRNA-valmisteille hätäkäyttöluvan myöntänyt Yhdysvaltain lääkeviranomainen FDA yrittivät yhdessä salata Pfizerin lupahakemuksen yhteydessä FDA:lle luovuttamat tutkimustulokset 75 vuodeksi (viite 61). Tuomioistuimeen tehty valitus johti kuitenkin määräykseen julkaista tulokset (viite 62), joista paljastui, että Pfizer ja FDA olivat tietoisia valmisteen runsaista terveyshaitoista, jotka olivat niin merkittäviä, että valmisteita ei olisi koskaan pitänyt päästää markkinoille. Kaikki tämä tieto on myös toimitettu kaikkien muiden maiden terveysviranomaisille, joten mm. THL:n on täytynyt olla siitä tietoinen ennen koronarokotusten alkua. Paljastusten seurauksena jo viisi Yhdysvaltain osavaltiota on haastanut Pfizerin oikeuteen koronarikoksista (viite 63).
Tässä vaiheessa on syytä pysähtyä kysymään, miten on mahdollista että:
- Kokeellinen valmiste, jonka tiedetään olevan tarpeeton ja tehoton, ja jonka valmistaja ei uskalla ottaa vastuuta sen aiheuttamista haitoista koska sen turvallisuutta ei ole tutkittu, voi päästä markkinoille
- Edellisestä tietoiset terveysviranomaiset valehtelevat valmisteen olevan tehokas ja turvallinen, ja tekevät kaikkensa saadakseen koko kansan rokotettavaksi
- Lääkevalvontaviranomaiset eivät tee elettäkään valmisteen poisvetämiseksi markkinoilta, vaikka rokotehaittarekistereihin on kirjattu ennennäkemätön määrä vakavia haittoja ja kuolemia
Koronahuijaus on niin läpinäkyvä, että kukaan asiaan perehtynyt ei voi olla sitä ymmärtämättä. Miten on siis mahdollista, että asiantuntijat eivät tulleet joukolla julkisuuteen tyrmäämään levitettyjä tiedevastaisia väitteitä? Syitä tähän ovat:
- Korruptio: Rokoteteollisuus lahjoi osan asiantuntijoita kertomaan valheita
- Aivopesu: Lahjotut korruptiomyönteiset rokotehaittadenialistit aivopesivät suuren joukon muita asiantuntijoita, jotka eivät itse tutustuneet asioihin vaan luottivat sokeasti kerrottuihin valheisiin
- Sensuuri: Valheita toitotettiin ihmisille massiivisella mediakampanjalla, ja tehokkaan sensuurin avulla hiljennettiin vastakkaiset näkemykset
- Pelko: Erityisesti lääkäreitä kiellettiin kritisoimasta koronatoimia ja kertomasta faktoja, ja kiellosta poikenneet leimattiin foliohattuisiksi salaliittoteoreetikoiksi ja heidän uransa pyrittiin tuhoamaan
Koronahuijauksen taustat alkavat olla hyvin selvillä, ja pelkästään Yhdysvalloissa on käynnissä yli 20.000 oikeudenkäyntiä koronatoimia vastaan (viite 64). Koronahuijaus on rokoteteollisuuteen massiivisesti sijoittaneeseen Bill Gatesiin ja hänen rokoteteollisuutta avokätisesti tukevaan säätiöönsä (Bill & Melinda Gates Foundation) vahvasti henkilöityvän länsimaisen ns. globalistieliitin masinoima operaatio ja harjoitus heidän tulevaisuudessa suunnittelemiaan uusia pandemioita varten, joiden toteuttamiseen tärkeimmät työkalut ovat Maailman Talousfoorumi (World Economic Forum, WEF) ja Maailman Terveysjärjestö (World Health Organisation, WHO).
Saksalaisen Klaus Schwabin vuonna 1971 perustaman ja johtaman WEF:n (viite 65) tavoitteena on ns. suuri nollaus eli Great Reset (viite 66), neljäs teollinen vallankumous (Fourth Industrial Revolution, viite 67) ja YK:n Agenda 2030 (viite 68). Näiden kaunopuheisten agendojen oikeisiin tavoitteisiin voi tutustua tarkemmin muualta (viite 69). WEF on ”kouluttanut” sätkynukeikseen suuren määrän poliitikkoja ja yritysjohtajia Young Global Leaders (YGL) -ohjelmassaan (viite 70), mm. täydellisellä ajoituksella korona-ajan pääministeriksi Suomessa nostetun Sanna Marinin sekä monien muiden maiden korona-aikaiset johtavat poliitikot. Globalistien tähtäimessä on ihmisten totalitaristinen kontrollointi digitaalisella henkilökortilla ja pankkijärjestelmällä, joiden avulla ihmiset voidaan mm. pakottaa ottamaan globalistieliitin kulloinkin vaatimat huuhaarokotteet. Koronapassien avulla valtiot saatiin pystyttämään tarvittavat järjestelmät, joilla tätä suunniteltua totalitarismia harjoiteltiin.
”Every politician with ties to World Economic Forum should be arrested on suspicion of high treason”
Itse asiassa Pfizerin ja Modernan mRNA-teknologiaan perustuvat ns. koronarokkotteet eivät ole rokotteita lainkaan vaan geeniterapioita (viite 71, viite 72, viite 73). Niiden väitettiin valheellisesti olevan rokotteita, koska geeniterapioiden hyväksymisprosessi on huomattavasti pidempi, ja niille ei voida myöntää sellaista hätäkäyttölupaa, jonka nojalla niitä voitaisiin ylipäätään antaa ihmisille nopealla aikataululla. Harva ihminen olisi myöskään suostunut piikitettäväksi, jos heille olisi kerrottu, että kyseessä oli rokotteen sijasta kokeellinen geeniterapia eli muuntogeeninen GMO-valmiste. Suomessa eduskunta hyväksyi 3.6.2021 geenitekniikkalain muuttamisen siten, että se mahdollisti koronarokotteiksi kutsuttujen kokeellisten GMO-valmisteiden antamisen kansalaisille (viite 74). Tämä tehtiin kaikessa hiljaisuudessa kansalaisilta salassa samalla kun mediassa kansan huomio kiinnitettiin pääministeri Sanna Marinin aamiaiskohuun (viite 75). Nuorempana lääketeollisuutta ja Pfizeria blogikirjoituksissaan voimakkaasti kritisoinut Marin (viite 76, viite 77) oli ehkä tajunnut millaisen huijauksen toteuttamiseen hänet oli valjastettu, jolloin valtamediaa hallitsevat globalistit näyttivät aamiaiskohun avulla, miten he voivat halutessaan tuhota hänen uransa, ellei hän tottele.
Yllä mainittu geenitekniikkalain muutosta koskeva poikkeusmenettely päättyi kaikessa hiljaisuudessa 26.3.2024 (viite 78). Tiedotteessa aiheesta todetaan varsin yksiselitteisesti ”COVID-19-pandemian aikana voimassa ollut poikkeus koronataudin hoitoon tai ehkäisyyn tarkoitettujen GMO-lääkkeiden lupamenettelylle on päättynyt EU:n alueella 6.3.2024.” Menettelystä käy yksiselitteisen selkeästi ilmi, että poliitikoilla ja terveysviranomaisilla oli koko ajan tiedossa, että koronarokotteiksi kutsutut valmisteet eivät olleet rokotteita vaan kokeellisia geeniterapioita eli muuntogeenisiä GMO-valmisteita, ja että tämä tieto salattiin kansalaisilta, joita pyrittiin perustuslakia ja ihmisoikeuksia törkeästi rikkomalla painostamaan ja kiristämään ottamaan kyseisiä valmisteita. Ottaen huomioon, että terveysviranomaisten ja poliitikkojen täytyi jo kyseisiä valmisteita hankkiessaan olla tietoisia myös siitä, että ne ovat tarpeettomia, tehottomia ja hengenvaarallisia, heidän toimintansa täyttää Suomen kansalaisiin tietoisesti kohdistetun kansanmurhan tunnusmerkit, ja vastaava kansanmurha toteutettiin samoilla keinoilla maailmanlaajuisesti globalistien ohjauksessa.
Globalistieliitti pyrkii luomaan yhteiskuntajärjestelmän, jota he kutsuvat nimellä ”Stakeholder Capitalism” (viite 79), joka on yhdistelmä fasismia ja kommunismia (viite 80). Fasismilla tarkoitetaan yhteiskuntajärjestelmää, jossa valtiot ja suuryritykset ovat liittoutuneet, kun taas kommunismilla tarkoitetaan yhteiskuntajärjestelmää, jossa valtio omistaa kaiken ja yksityisomistuksia ei sallita. Stakeholder Capitalismissa globalistieliitti omistaa suuryritykset, joiden tuotteita valtiot ostavat valtavia määriä veronmaksajien rahoilla, mikä täyttää fasismin tunnusmerkit. Stakeholder Capitalismin kommunismissa suuryritykset rosvoavat valtioiden ja veronmaksajien rahat itselleen, pienemmät yritykset tuhotaan ja suuryritysten lisäksi muita yksityisomistuksia ei sallita.
Koronapandemia oli globalistien harjoitus Stakeholder Capitalismin toteuttamiseen. Valepandemialla kansa peloteltiin hyväksymään turhat ja haitalliset yhteiskunnan rajoitustoimet ja valerokotteet. Rajoitustoimet aiheuttivat runsaasti haittaa pienemmille yrityksille, virusten leviämisen ehkäisemiseen tehottomaksi tiedetyillä kasvomaskeilla lisättiin hysteriaa ja pelon tunnetta, valtiot ostivat kansalaisilleen valtavan määrän valerokotteita, ja koronapassia käytettiin kiristämään kansalaisia ottamaan valerokotteet ehtona rajoitusten välttämiselle. Hoitohenkilökunta käytännössä pakotettiin ottamaan valerokotteet tartuntatautilain 48a-pykälän avulla, mikä johti joukkopakoon alalla, kun valtavan määrän rokotehaittoja nähneet hoitajat eivät suostuneet vaarantamaan terveyttään.
Koronapassi ja 48a saatettiin väkisin voimaan, vaikka niistä päätettäessä oli yleisesti tiedossa, että valerokotteet eivät estä saamasta tartuntaa, levittämästä virusta edelleen ja tartuttamasta muita. Pfizer myönsi Euroopan Parlamentin kuulustelussa lokakuussa 2022, että mRNA-valmisteiden tullessa markkinoille asiaa ei oltu edes tutkittu (viite 81). Euroopan lääkeviranomainen EMA myönsi myös marraskuussa 2023 asiaa tiedustelleille europarlamentaarikoille, että koronarokotteille oli myönnetty lupa ainoastaan yksittäisten henkilöiden suojeluun, ja että niitä ei voida käyttää koronaviruksen leviämisen estämiseen (viite 82, europarlamentaarikoiden virallinen lehdistötiedote aiheesta viitteessä 83). Suomessakin asiantuntijat olivat asiasta perillä vielä ennen rokotusten aloittamista loppuvuonna 2020 (viite 84), mutta jossain vaiheessa rokotehuumaa/pelkoa asia ”unohtui” päättäjiltä. Koronapassille ja 48a:lle ei siis ollut missään vaiheessa mitään lääketieteellistä perustetta, vaan kyseessä oli puhtaasti totalitaristisen terveysfasismin toteuttaminen, jolla kansalaiset pyrittiin pakottamaan ottamaan tarpeettomaksi, tehottomaksi ja haitalliseksi tiedetyt valerokotteet.
Koronapassi ja tartuntatautilain 48a-pykälä olivat törkeitä ihmisoikeusrikkomuksia. Kun valtio hankkii kansainväliseltä suuryritykseltä turhia ja hengenvaarallisia tuotteita ja tekee kaikkensa pakottaakseen kansalaisensa käyttämään niitä, ollaan totalitaristisen äärioikeistolaisen fasismin ytimessä (viite 85). Tätä fasismia ajoi Suomessa voimakkaimmin SDP Krista Kiurun ja globalistien YGL-koulutetun Sanna Marinin johdolla. Rokottamattomien törkeä leimaaminen ja ajojahti valtamediassa sai ennennäkemättömät rasistiset mittasuhteet, jotka muistuttivat Natsi-Saksan aikaisia juutalaisvainoja (viite 86).
Onko siis niin, että Suomen johtava äärioikeistolainen fasistipuolue on SDP? Nykyisistä eduskuntapuolueista koronapassia vastustivat ainoastaan Kristillisdemokraatit ja usein äärioikeistolaisiksi leimatut Perussuomalaiset, jotka siis nousivat ainoina puolueina vastustamaan fasismia. Onkin varsin mielenkiintoista, että ympäri Eurooppaa nimenomaan fasistisia koronatoimia vastustaneet puolueet, kuten Saksan AfD ja Itävallan FPÖ, on leimattu äärioikeistolaisiksi fasisteiksi muiden, fasistisia koronatoimia ajaneiden puolueiden toimesta. Ilmeisesti oikeat fasistit käyttävät laajalti ”hyökkäys on paras puolustus” -taktiikkaa peitelläkseen omia fasistisia tarkoitusperiään. Kaikkien onkin nyt hyvä ymmärtää, että valtamediassa ”äärioikeistolainen” tarkoittaa nykyään fasismia vastustavia puolueita, ja kaikki muut puolueet paitsi ”äärioikeistolaiset” kannattavat fasismia.
Kaikissa länsimaissa, Suomi mukaanlukien, on käynnissä massiivinen globalistien masinoima operaatio, jossa johtavat poliittiset puolueet on valjastettu ajamaan toimia, jotka tähtäävät Stakeholder Capitalism -yhteiskuntajärjestelmän luomiseen. Tällä fasismia ja kommunismia yhdistävällä politiikalla on kolme pääasiallista tavoitetta: kansanterveyden, kansantalouden ja valtioiden itsenäisyyden tuhoaminen. Näitä tavoitteita ajoi Suomessa ansiokkaasti ja onnistuneesti Sanna Marinin hallitus, josta Marin sai myöhemmin palkinnoksi asiantuntijatoimen toisen globalistien sätkynuken Tony Blairin organisaatiossa (viite 87, viite 88).
Kansalaisaktivisti Mika Vauhkala haastoi Suomen valtion ja Fazerin oikeuteen koronapassista, koska hän ei saanut joulukuussa 2021 Fazerin kahvilassa aamiaista ilman koronapassia (viite 89). Vauhkalan toiminta on esimerkillistä kansalaisaktiivisuutta, jossa pyritään saamaan valtio ja Fazer vastuuseen kansalaisten perustuslaillisia oikeuksia törkeillä ihmisoikeusrikkomuksilla loukkaavasta koronapassista. Tämä on tärkeää myös siksi, että valtion ja Fazerin tuomitsemisen myötä estettäisiin vastaavat ihmisoikeusloukkaukset globalistien suunnittelemissa uusissa valepandemioissa. Vauhkala ja hänen tiiminsä esittivät käräjäoikeudelle laajat kiistattomat todisteet, mutta käräjäoikeus kuitenkin tuomitsi valtion ja Fazerin voittajaksi, ja hovioikeus hylkäsi Vauhkalan valituksen.
Vauhkalan käräjäoikeudelle ja hovioikeudelle esittämät todisteet olivat niin selkeät, että kukaan asiaan perehtynyt ei voi olla niitä ymmärtämättä. Vauhkalan kokema törkeä oikeusmurha vahvistaa ilman mitään epäilyjä sen, että sekä asiaa käsitelleet käräjäoikeus että hovioikeus ovat globalistien hallinnassa, ja Suomi ei ole enää oikeusvaltio. Fazer-konsernissa, jonka oletan joutuneen tapauksessa valtion painostuksen alaiseksi, ei myöskään voida mitenkään olla ymmärtämättä asiaa. Toivoisinkin Fazerilta julkista ulostuloa, jossa he myöntävät koronapassilla valtion painostuksesta toteutetut ihmisoikeusrikkomukset ja pyytävät niitä anteeksi globalistien hallitsemien oikeusasteiden päätöksestä huolimatta. Henkilökohtainen mielipiteeni on, että kaikkien koronapassia vaatineiden tahojen tulisi myöntää syyllistyneensä vakaviin ihmisoikeusrikkomuksiin ja pyytää niitä julkisesti anteeksi, ja ne jotka eivät näin tee tulisi myöhemmin saattaa rikosoikeudelliseen vastuuseen.
Viimeiset vuodet ovat vahvistaneet, että globalistien kaappaaman WHO:n yhtenä pääasiallisena tavoitteena on rokotemyynnin edistäminen keinoja kaihtamatta. Bill & Melinda Gates -säätiöstä on tullut WHO:n merkittävä rahoittaja, ja sen myötä WHO:sta on tullut globalistieliitin kaappaama työkalu. WHO muutti hieman ennen koronapandemian julistamista pandemian määritelmää sellaiseksi, että se voi jatkossa julistaa pandemioita mielivaltaisesti (viite 90). Globalisteilla lienee ollut hauskaa kun he ovat keksineet valepandemiajulistuksilleen virallisen nimen ”Public Health Emergency of International Concern”, joka lyhennetään ”PHEIC” ja lausutaan kuten ”FAKE”.
WHO on viime vuosina valmistellut kaikessa hiljaisuudessa ns. Pandemic Treaty -sopimusta (viite 91) ja muutoksia kansainväliseen terveyssäännöstöön (International Health Regulations, IHR, viite 92), tavoitteenaan että kaikki WHO:n jäsenvaltiot luovuttavat tulevissa valepandemioissa kaiken määräysvallan WHO:lle. Mikäli tämä toteutuu, WHO voi mielivaltaisesti julistamissaan uusissa valepandemioissa määrätä jäsenvaltionsa toteuttamaan haluamiaan toimia, kuten yhteiskunnan rajoitustoimia, maski- ja rokotuspakkoja, ja määrätä rokotuksista kieltäytyminen rangaistavaksi. Toteutuessaan tämä asettaisi WHO:n diktatoriseen asemaan toteuttamaan vapaasti totalitaristista fasisitista terveystyranniaa, antaen kaiken vallan rokoteteollisuuden jättivoittojen kautta entisestään rikastuvalle globalistieliitille.
Kaunopuheisista valheista huolimatta Pandemic Treaty ja IHR-muutokset ovat kokeneet sellaista vastustusta erityisesti Afrikan maiden taholta, että niiden voimaantulo on merkittävästi viivästynyt (viite 93). Asian korjaamiseksi WHO julisti 14.8.2024 apinarokkovalepandemian (viite 94), jonka tarkoituksena lienee Afrikan maiden kiristäminen hyväksymään Pandemic Treaty ja IHR-muutokset. Tämä ei kuitenkaan ole edelleenkään johtanut globalistien haluamaan tulokseen, mikä viittaa siihen, että koronan tapaiset huijaukset eivät mene enää läpi ilman Pandemic Treatya ja IHR-muutoksia.
WHO:lla on pitkä lista valmiita suunnitelmia uusista valepandemioista (viite 95). Listan kärjessä on lintuinfluenssa (viite 96), jota on valmisteltu jo pitkään, ja jolle on jo valmiiksi kehitetty tarpeettomiksi, tehottomiksi ja hengenvaarallisiksi tiedetyt rokotteet, joita huijattiin annettavaksi ensimmäisenä sinisilmäisille Suomessa (viite 97, viite 98). Muita suunnitteilla olevia valepandemioita ovat ainakin Marburg-virus (viite 99) sekä enterorokko (SEERS, viite 100). Lintuinfluenssan väitetään olevan vakava uhka ihmisille, vaikka sen ei ole koskaan todettu tarttuneen ihmisiin, eikä ole mitään perusteita sille, että se voisi ihmiseen tarttuvaksi muuttua. Lintuinfluenssan pelossa on Suomessakin lahdattu turkiseläimiä, joiden väitetään perusteitta voivan toimia välittäjinä lintuinfluenssaviruksen tarttumisessa ihmiseen (viite 101). Tulevissa valepandemioissa on tosin vaarana se, että globalistit olisivat tällä kertaa, toisin kuin koronaviruksen osalta aiemmin, onnistuneet kehittämään laboratoriossa oikeasti vaarallisen viruksen, jonka he päästäisivät irti sen jälkeen kun WHO:n Pandemic Treaty ja IHR-muutokset on hyväksytty. Mikäli näin kävisi, mitään rokotteita ei kuitenkaan tarvita, sillä esim. ivermektiini tiedetään hyvin tehokkaaksi lääkkeeksi RNA- ja DNA-viruksia vastaan (viite 102).
Donald Trumpin murskavoitto Yhdysvaltain presidentinvaaleissa on selkeä osoitus siitä, että Yhdysvalloissa kansa on laajalti herännyt ja saanut tarpeekseen globalistien kaappaaman Bidenin/Harrisin hallinnon fasismiin ja kommunismiin tähtäävästä politiikasta. Kyseessä on ehkä kaikkien aikojen tärkein vaalitulos koko maailman, myös Suomen kannalta, mikäli Trump pitää vaalilupauksensa. Trump on mm. luvannut varsin suorasanaisesti saattaa globalistit ja kaikki heidän talutusnuorassaan koronatoimiin osallistuneet tahot vastuuseen teoistaan (viite 103, viite 104). Näitä tahoja ovat mm. poliitikot, terveysviranomaiset, lääkevalvontaviranomaiset, lääkärit, asiantuntijat, media ja sen toimittajat, sekä erilaisia sulku- ja rajoitustoimia ajaneet ja toteuttaneet tahot. Trump on jo nimittänyt vaalilupauksensa mukaisesti Yhdysvaltain terveysministeriksi Robert F. Kennedyn (viite 105), joka on puhunut paljon julkisuudessa koronahuijauksesta. Kennedy onkin jo ehtinyt nimityksensä jälkeen antamaan julkisuudessa lausuntoja, joiden mukaan hän tulee siivoamaan Yhdysvaltain terveysviranomaiset täydellisesti rokoteteollisuuden korruptoimista huijareista, ja myös rokoteteollisuus tullaan asettamaan rikosoikeudelliseen vastuuseen (viite 106). Trumpin ja Kennedyn virkakausi alkaa 20.1.2025, ja radikaaleja toimenpiteitä on odotettavissa tämän jälkeen nopealla aikataululla.
Trump ja Kennedy ovat äärimmäisen vakava uhka globalisteille, joiden tulevaisuus näyttää muutenkin hyvin huonolta, viitaten tuoreeseen lumipalloilmiöön, jossa Ranskan (viite 107) ja Saksan (viite 108) globalistihallitukset kaatuivat ja yksi pahimmista globalistien sätkynukeista, WEF:n YGL-koulutettu Kanadan pääministeri Justin Trudeau joutui eroamaan (viite 109). Näyttääkin siltä, että globalistieliitin taru on tulossa tiensä päähän, jolloin heidän vaikutusvaltansa eri oikeusasteissa heikkenee olennaisesti, mistä tulee seuraamaan valtava vyöry tuomioita käynnissä olevissa kymmenissä tuhansissa oikeudenkäynneissä koronatoimia vastaan (viite 63). Tämän seurauksena ennustan, että vielä Trumpin nelivuotisen presidenttikauden aikana järjestetään kansainvälinen ”Nuremberg 2” -sotaoikeudenkäynti, jossa globalistit ja kaikki heidän talutusnuorassaan olevat, myös Suomessa koronatoimia ajaneet ja toteuttaneet tahot tullaan saattamaan rikosoikeudelliseen vastuuseen.
Alla vielä lista muutamista tärkeistä toimenpiteistä koronatoimia vastaan, joita ei ole aiemmin tässä kirjoituksessa mainittu:
- Marraskuussa 2024 tietovuoto paljasti Robert Koch Instituutin (Saksan lääkeviranomainen) pelotelleen kansaa tietoisesti valheilla, ja tiedevastaisten koronatoimien tavoitteena oli kansalaisten piikittäminen tarpeettomilla, turhilla ja haitallisiksi tiedetyillä valerokotteilla (viite 110). Kaikki merkit viittaavat siihen, että toiminta oli globalistien tarkkaan koordinoimaa ja toteutettiin samanlaisena myös muissa maissa, Suomi mukaanlukien.
- Julkisuuteen vuodetussa EU:n ja Pfizerin allekirjoittamassa sopimuksessa Pfizer myönsi, että mRNA-valmisteen tehoa ja turvallisuutta ei ole asianmukaisesti tutkittu, minkä vuoksi Pfizerille myönnettiin sopimuksessa täysi vastuuvapaus (viite 111, sivu 24, kohta I.12.1 sekä sivu 48, kohta 4). Kansalaisille kuitenkin valehdeltiin valmisteen olevan tehokas ja turvallinen.
”Do the covid vaccines provide immunity? Yes, but only for the manufacturers”
- Euroopan komission puheenjohtajan Ursula von der Leyenin Pfizerin kanssa salaa neuvottelemasta koronarokotesopimuksesta on tehty kaksi erillistä rikosilmoitusta (viite 112, viite 113, viite 114).
- Swissmedicistä (Sveitsin lääkeviranomainen) on tehty rikosilmoitus mm. koronarokotteilla tietoisesti toteutetusta kansanmurhasta (“multiple intentional homicide”), laajan huolellisesti kootun näytön asiassa vaikuttaessa hyvin vahvalta (viite 115).
- Bill Gatesistä on tehty rikosilmoitus Hollannissa koronahuijauksesta ja koronarokotteiden aiheuttamista haitoista, ja oikeuden päätöksellä 16.10.2024 hän joutuu vastaamaan syytteisiin (viite 116).
- Kansainvälinen asiantuntijaryhmä ”North Group” vaatii 25.11.2024 eri maiden hallituksille lähettämässään kirjeessä koronarokotusten välitöntä keskeyttämistä (viite 117)
- Pfizerin kliinisen tutkimuksen alihankkijan Ventavian työntekijä Brook Jackson on tehnyt rikosilmoituksen Pfizeria vastaan tutkimustulosten vääristelystä (viite 118). Oikeudenkäynti on edelleen kesken, ja sen edistymistä voi seurata Jacksonin nettisivuilla, jonne on koottu kaikki tapaukseen liittyvät asiakirjat (viite 119).
Juoni tihenee, ja seuraavia paljastuksia ja tuomiopäivää odotellessa vielä mielenkiintoinen väite hautumaan mielen sopukoihin. Pitkän linjan lääkekehitysasiantuntija Sasha Latypova väittää, että kaikki valekoronarokotteet on tehty äärifasistisessa operaatiossa Yhdysvaltain puolustusministeriön (Department of Defence, DoD) toimeksiannosta, että ne on kehitetty alunperin ihmiselle haitallisiksi biologisiksi aseiksi joita on salaa testattu maailman väestöllä valepandemian varjolla (viite 120). Latypovan näyttö asiassa vaikuttaa vakuuttavalta, ottaen huomioon myös sen, että Yhdysvalloissa on aiemminkin tehty epäeettisiä testauksia mm. testaamalla ihmisillä salaa biologisia aseita (viite 121). Väitteet siitä, että koronarokotteet on kehitetty biologiseksi aseeksi esitti ensimmäisen kerran patenttiasiantuntija David Martin jo kesällä 2021 varsin vakuuttavien todisteiden kera (viite 4). Tämä mitä hulluimmalta foliohattujen salaliittoteorialta kuulostava väite selittäisi sen, miksi terveys- ja lääkevalvontaviranomaiset eivät ole puuttuneet koronahuijaukseen – niillä ei ole siihen valtuuksia koska kyseessä ei ole tavanomainen lääke/rokotekehitys vaan sotilasoperaatio, jossa testataan maailman väestöllä tarkoituksella hengenvaaralliseksi kehitettyä biologista asetta.
Kliinisen psykologian professori Mattias Desmetin mukaan koronahuijaus on toteutettu taitavalla psykologisella mediakampanjalla, jonka avulla ihmiset on vaivutettu ns. massapsykoosiin (viite 122). Nyt on viimeistään aika kaikkien tunnustaa tosiasiat ja ravistella itsensä hereille.
”It is much easier to fool people than to convince them that they have been fooled”
Demokratian ja vapauden puolesta, fasismia ja kommunismia vastaan
Viitteet:
- Woo et al 2010 Coronavirus genomics and bioinformatics analysis
- Doshi 2020 Covid-19: Do many people have pre-existing immunity?
- Patenttien jälki – pandemia rahastusoperaationa
- David Martin: Patenttitiedot paljastavat biologisen aseen
- Patent US7279327 Methods for producing recombinant coronavirus
- Korona sittenkin laboratoriosta
- Axfors and Ioannidis 2021 Infection fatality rate of covid-19
- Tilastokeskus kuolemansyyt 2020
- Paul Lillrank: Miksi meillä ei ole pandemiaa
- STM:n luokituksen mukaan korona ei ole pandemia
- TM Shokeeraava korrelaatio koronakuolemien ja ylipainon välillä
- Borsche et al. 2021 Covid-19 and vitamin D
- James Roguski: Honoring Kary Mullis
- Corman Drosten Review Report
- Bullard et al 2020 Predicting infectious SARS-CoV-2 from diagnostic samples
- Jaafar et al. 2021 Correlation between PCR and cell cultures
- Surkova et al. 2020 False-positive Covid-19 results: hidden problems and costs
- WHO information notice SARS-CoV-2 PCR
- Griffin 2020 Asymptomatic cases may not be infectious, Wuhan study indicates
- Madewell et al 2020 Household transmission of SARS-CoV-2
- Are face masks effective? The evidence
- Face masks: It is time to follow the science
- FIN unmasked 2021: Selvitys kasvosuojainten ja hygieniakäytäntöjen tehosta
- Professori: Käsidesin liiallinen käyttö on terveydelle haitallista
- America’s Frontline Doctors: Lockdowns do not work
- Covid-19 and vitamin D publications
- Tiedekooste D-vitamiini tehokas koronaa vastaan
- Meri 2015 Duodecim: Lääketieteen Nobel ivermektiinille
- Ivermectin reduces covid-19 risk, 105 studies
- Ivermektiinin hoito-ohjeet koronaan
- WHO guidance Corticosteroids for covid-19
- Steroidit vähentävät koronakuolemia
- Valtioneuvoston asetus koronatuista
- Cabalin kaatuminen osa 22 – Suurin lääketieteellinen huijaus: Covid-19
- Silva 2021 Kidney damage and remdesivir
- Ghosh 2020 Liver injury and remdesivir
- Ventilator-associated pneumonia
- Covid-19 vaccine provider incentive program
- Covid vaccine bribery scheme in USA
- Olliaro et al. 2021 The Lancet: Covid vaccine efficacy ARR and RRR
- Koronarokotteiden todellinen teho vain prosentin luokkaa
- Subramanian and Kumar 2021 Increases in covid-19 are unrelated to vaccination
- Doshi 2020 BMJ: Current trials are not designed to show if covid vaccines save lives
- Doctors For COVID Ethics 2023 mRNA vaccine toxicity
- Scientific publications of COVID vaccine toxicity
- EU 2 miljoonan koronapiikkihaitan raja lähestyy 07-2021
- Beattie 2021 Analysis of Vaccine Administration on Deaths and Cases Associated with COVID-19
- Rancourt et al. 2024 All-cause mortality in 125 countries during COVID
- CHD 2023 Increase in miscarriages and stillbirths linked to COVID shots
- Decline of live births in Europe
- THL suosittelee iäkkäille koronarokotuksia syksylle 2023
- Korona- ja influenssarokotukset syksyllä 2023
- IS 29.11.2023 Syksyn korona-aalto osui iäkkäämmin voimakkaammin kuin koskaan ennen
- IS 8.12.2023 Suomessa kuoli viikossa ennätysmäärä ihmisiä
- Robert Malone When is mRNA not really mRNA
- Rubio-Casillas et al. 2024 Pseudouridine and cancer
- McKernan et al. 2023 Moderna and Pfizer mRNA vaccines contain DNA
- Speicher et al. 2023 DNA fragments in Pfizer and Moderna mRNA vaccines
- Alden et al. 2022 Pfizer-BioNTech covid vaccine transcribes into DNA
- Koronapiikkien ainesosat, ei ihmiskäyttöön
- FDA määrätty luovuttamaan Pfizerin k-piikin turvallisuusdata
- US District Court Order to FDA
- Five US states launch massive lawsuits against Pfizer
- Steve Kirsch over 20,000 covid legal cases, new lawyers needed
- Wikipedia Maailman Talousfoorumi
- WEF The Great Reset
- WEF The Fourth Industrial Revolution
- UN Agenda 2030
- Secret plans of World Economic Forum
- WEF Young Global Leaders
- Banoun 2023 mRNA vaccines are gene therapies
- Bayer CEO mRNA vaccines are gene therapies
- Court ruling mRNA jabs are not vaccines
- Eduskunta hyväksyi geenitekniikkalain muuttamisen 06-2021
- HS 31.5.2021 Pääministerin ateriakohu
- Sanna Marin blogi Raharikas lääketeollisuus
- Sanna Marin blogi Tampereen kaupunki Pfizerin lahjonnan kohde
- Geenitekniikkalautakunta Covid rokotteiden poikkeusmenettely GMO-valmisteille päättynyt
- WEF Stakeholder Capitalism
- Stakeholder Communism
- Pfizer EU hearing
- EMA response letter to European Parliament
- European Parliament press conference on the letter from EMA
- IS 10.11.2020: Koronarokote suojaa vain saajaa, ei estä tartuntoja
- Bell 2024 Pandemic preparedness and the road to international fascism
- Koronatoimissa kaikuja Natsi-Saksasta
- WEF Tony Blair Institute for Global Change
- YLE Sanna Marin Tony Blair -Instituutti
- Mika Vauhkala Case Covidpass
- Vapauspassi ja WHOn uusmääritelmät
- Time to stop WHO Pandemic Treaty 22.5.2024
- James Roguski 9.3.2023 Silence procedure for amendments to IHR
- Kantomedia 31.5.2024 WHOn pandemiasopimus kaatui
- IL 14.8.2024 WHO julisti apinarokkopandemian
- WHO plans for 10 years of pandemics
- McCullough Foundation 18.6.2024 Bird Flu, separating fact from fiction
- CHD 27.6.2024 Finland to offer bird flu vaccine
- PSL 21.8.2024 WHO pelottelee uusilla taudeilla ja markkinoi vaarallisia rokotteita
- IL 4.10.2024 Marburg epidemia Afrikassa
- Catastrophic Contagion SEERS pandemic 2025 simulation
- PSL 28.9.2023 Lintuinfluenssa ei ole koskaan tarttunut ihmiseen
- FLCCC Alliance The remarkable story of ivermectin
- Donald Trump Covid tyrants – We will not comply
- Trump outlines plan to destroy deep state
- YLE 14.11.2024 Trump nimitti RFKn terveysministeriksi
- NY Post 11.11.2024 RFK suggests 600 people fired from NIH
- YLE 4.12.2024 Ranskan hallitus kaatui
- IS 16.12.2024 Saksan hallitus kaatui
- IS 6.1.2025 Justin Trudeau eroaa
- Stunning revelations in massive Robert Koch Institute leak 9.11.2024
- EU Pfizer covid vaccine agreement
- Politico 1.4.2024 Pfizergate EU covid vaccine scandal
- IS 17.7.2024 EU komissio ei olisi saanut salata koronarokotesopimusten tietoja
- Euronews 15.11.2024 Pfizergate battle kicks off in European Court
- Criminal complaint against Swissmedic 7.2.2024
- Bill Gates loses lawsuit and will appear in Dutch court 20.4.2024
- North Group info
- Thacker 2021 BMJ Researcher blows the whistle on data integrity issues in Pfizer’s vaccine trial
- Brook Jackson Case documents
- Sasha Latypova Jan 2023 Covid-10 countermeasures, Intention to harm
- Unethical human experimentation in USA
- Mattias Desmet Miksi niin monet uskovat yhä viralliseen narratiiviin